The retinal pigment epithelium (RPE) contains not only the usual cellular organelles found in most cells (such as the nucleus, Golgi apparatus, smooth and rough endoplasmic reticulum, and mitochondria) but also melanin granules and phagosomes, which signify its two important functions. The RPE is rich in small peroxisomes, suggesting its active involvement in detoxifying a large quantity of free radicals and oxidized lipids in a highly oxidative and well-lit environment. Despite being a monolayer structure, the RPE performs a variety of complex biochemical functions, including the transport and metabolism of vitamin A, drug detoxification, the synthesis of melanin and extracellular matrix, and the selective transfer of nutrients and metabolites between the outer retina and the choroid. It also phagocytoses and digests the shed membrane discs of photoreceptor outer segments and serves as a pigment barrier to maintain the environment necessary for photoreceptor activity. Tight junctions between RPE cells prevent macromolecules in the fluid leaking from choroidal blood vessels from entering the retina, acting as part of the blood-retinal barrier (together with Bruch's membrane, forming the retina-choroid barrier). Biochemically, the RPE exhibits dynamic complexity, accommodating its active metabolism, specialized phagocytic functions, and its role as a biologic filter for the sensory retina. These functions impose significant energy demands, which are met by the enzymatic activity of three major biochemical pathways in RPE cells: glycolysis, the tricarboxylic acid cycle, and the pentose phosphate pathway.
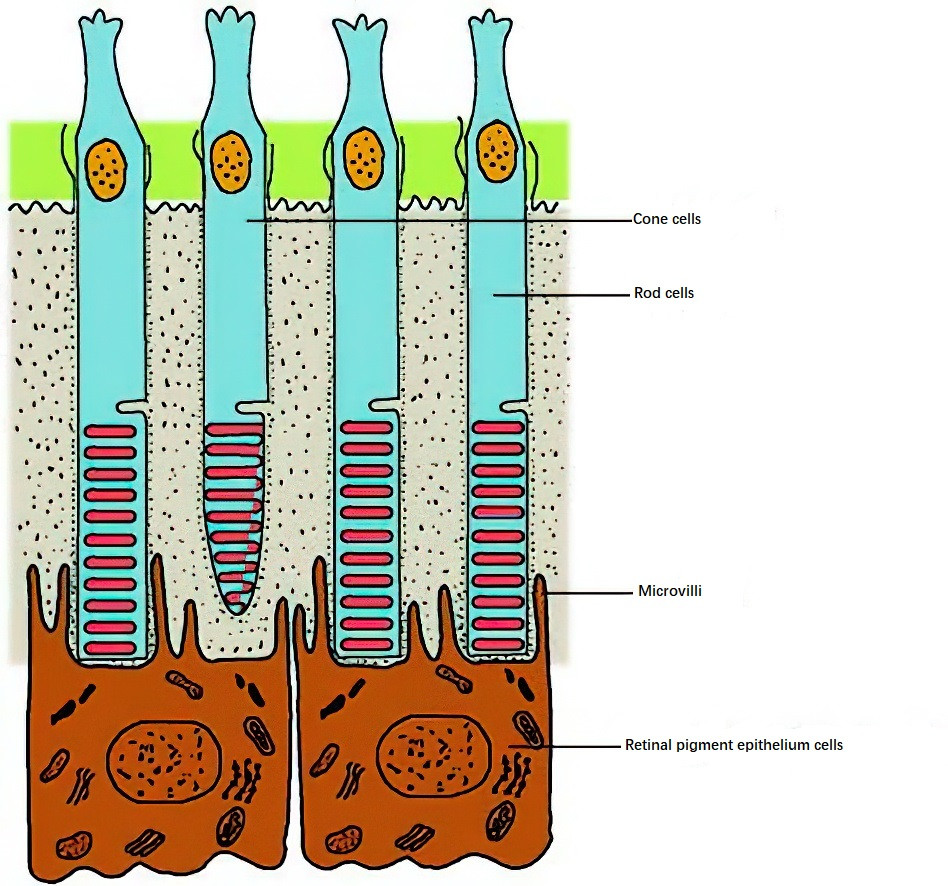
Figure 1 Diagram depicting the relationship between the retinal pigment epithelium and photoreceptors
Additionally, the RPE promotes anatomical adhesion between the retina and the choroid. Glial cells in the retina, particularly Müller cells, span the entire sensory retina, with fibers extending longitudinally from the outer limiting membrane to the inner limiting membrane, providing structural support and metabolic nutrition to the retina.
Visual information is transformed into neural impulses in the retina and transmitted through three levels of neurons: photoreceptors, bipolar cells, and ganglion cells. The axons of ganglion cells form the nerve fibers that carry visual information along the visual pathway to the lateral geniculate nucleus (the fourth-order neuron), from which it is transmitted to the visual cortex for the perception of vision.
Photoreceptors are the first-order neurons in the retina and are categorized into rods and cones. Rod cells are responsible for dim-light (scotopic) vision and achromatic vision, while cone cells mediate bright-light (photopic) vision and color perception. Approximately 7 million cone cells are concentrated mainly in the macula. The central fovea contains only cone cells, and the neural pathways here exhibit single-line connectivity, resulting in exceptionally sharp vision. Beyond the central fovea, the density of cone cells decreases significantly. As a result, macular lesions lead to a noticeable decline in visual acuity. Rod cells are absent in the central fovea but start appearing at approximately 0.13 mm from the fovea and increase in number, reaching their maximum density around 5 mm, after which their number gradually decreases toward the periphery. Damage to peripheral retinal areas can cause rod cell loss, leading to night blindness. The optic disc, where the nerve fibers converge to form the optic nerve, lacks photoreceptor cells and thus has no visual function, presenting as a physiological blind spot in the visual field.
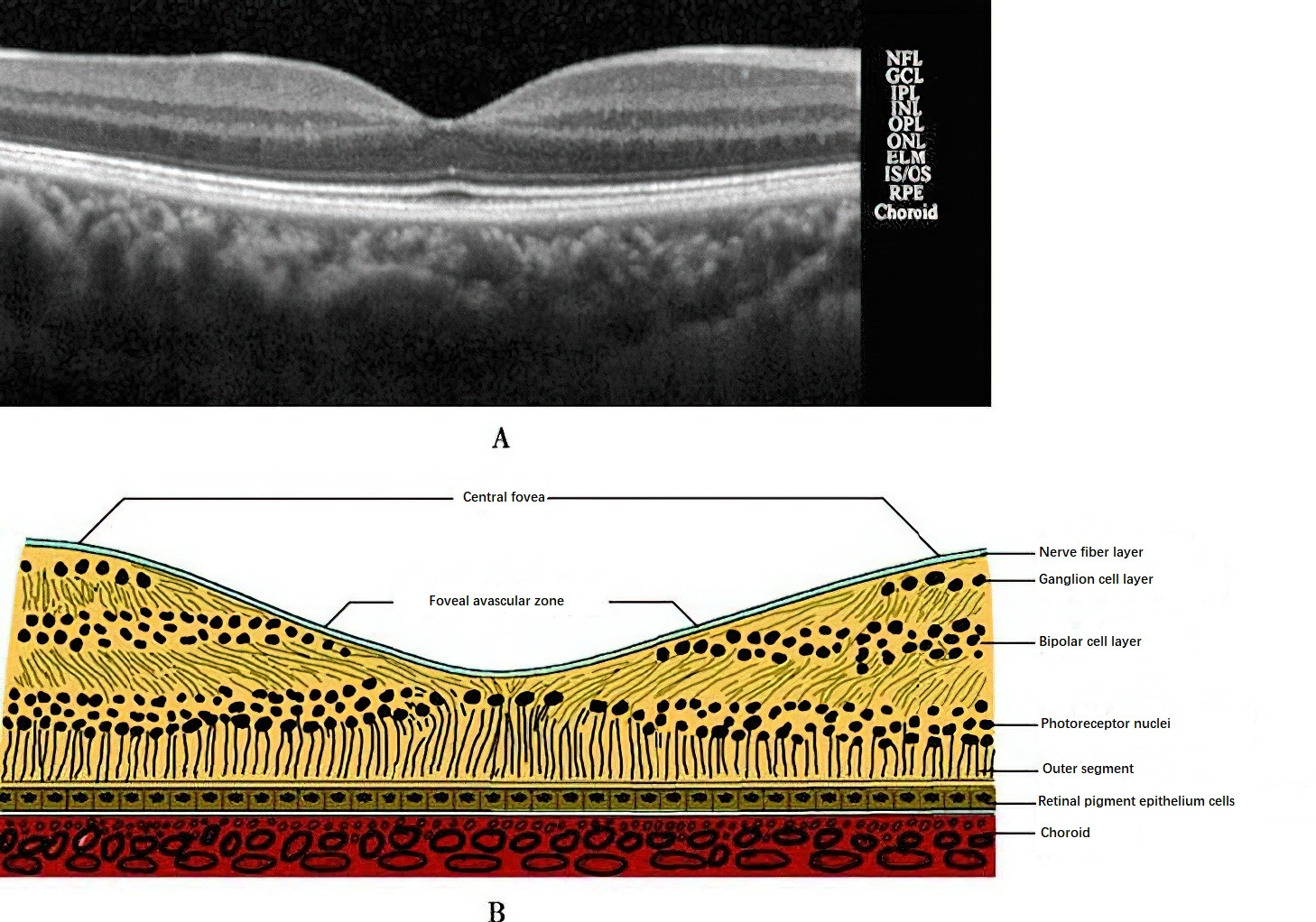
Figure 2 Structure of the macula
Each photoreceptor's outer segment contains only one type of photopigment. Rod cells contain rhodopsin, which is a combination of 11-cis-retinal and opsin. In darkness, the regeneration of rhodopsin increases the retina's sensitivity to dim light. Cone cells contain three types of photopigments for color vision: iodopsin, erythrolabe, and cyanolabe. These photopigments, synthesized from other forms of vitamin A aldehyde and opsin, mediate color perception in response to light. Therefore, color vision is a function of cone cells under well-lit conditions. The macula exhibits the highest sensitivity to color, which decreases with increasing distance from the macula. The peripheral retina has minimal color perception, consistent with the distribution of cone cells.
Several theories exist to explain color vision, with the most widely accepted at the retinal level being the Young-Helmholtz trichromatic theory. Normal color vision involves three photopigments in cone cells, each responsive to light of specific wavelengths: long (570 nm, corresponding to red), medium (540 nm, corresponding to green), and short (440 nm, corresponding to blue). Each photopigment elicits its strongest response to its corresponding primary color of light, with weaker responses to other colors. For example, red light strongly excites the pigment sensitive to red, moderately excites the green-sensitive pigment, and weakly excites the blue-sensitive pigment, thus producing a full spectrum of color perception. The absence of one type of photopigment in cone cells can result in color vision deficiencies.